Thème de recherche n°1 : Contrôle de la biosynthèse de l’appareil traductionnel et le rôle de la stabilité/maturation de l’ARNm pour l’expression génétique
Les gènes structuraux et les protéines qu’ils codent sont souvent bien conservés entre les différentes espèces bactériennes (i.e. des bactéries Gram positives et Gram négatives). En revanche, et de façon surprenante, les mécanismes qui contrôlent l’expression de ces gènes peuvent être très différents. Ceci est remarquable lorsque l’on compare les deux organismes modèles Escherichia coli (Gram-) et Bacillus subtilis (Gram+) distants de plus de deux milliards d’années d’évolution. Afin de mieux cerner les différentes stratégies utilisées pour moduler l’expression génétique chez les procaryotes il est donc important d’étudier plus d’un organisme de façon approfondie. Nous nous intéressons particulièrement aux mécanismes co- et post- transcriptionnels qui contrôlent la synthèse des composants de l’appareil traductionnel chez B. subtilis et particulièrement les mécanismes d’antiterminaison transcriptionnelle et de maturation/dégradation des ARNm. Récemment, nous avons étendu nos études à la caractérisation d’une communauté de B. subtilis en migration (« swarming »), un exemple de comportement social qui permet une analyse de l’expression génique en cellule unique.
Contrôle de l’expression de protéines de l’appareil traductionnel
La synthèse de l’appareil traductionnel (ribosomes, aminoacyl-ARNt synthétases, facteurs de traduction etc.) consomme plus de la moitié de l’énergie cellulaire et doit donc être contrôlée efficacement. Chez B. subtilis et les bactéries à Gram positif en général, nos connaissances de l’expression des protéines ribosomales sont encore assez rudimentaires notamment en ce qui concerne la stratégie utilisée pour coordonner leur synthèse avec celle de l’ARNr. Nous avons élucidé, au niveau moléculaire, le premier mécanisme qui contrôle l’expression d’une protéine ribosomale dans une bactérie Gram positif (l’opéron infC-rpmI-rplT chez B. subtilis). De façon surprenante, les deux organismes utilisent la même molécule effectrice (la protéine ribosomale L20) et du mimétisme moléculaire mais dans des contextes très différents : antiterminaison transcriptionnelle chez B. subtilis et autorépression traductionnelle chez E. coli. Ceci démontre que le mimétisme moléculaire, même s’il ne repose pour le moment que sur quelques exemples, est un concept très puissant capable d’être intégré dans des mécanismes de contrôle très divers. Le contrôle de la dégradation d’un ARNm peut être un moyen très efficace pour contrôler l’expression génique. Il constitue un outil puissant pour une adaptation rapide aux changements nutritionnels et environnementaux et permet à la cellule de contrôler de manière efficace la stœchiométrie de protéines codées par un opéron.
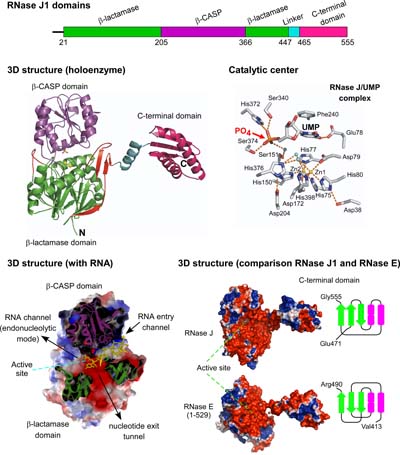
La RNase J1 est une ribonucléase à double activité, endo- et 5’-3’ exonucléolytique. Nous avons résolu la structure 3D de cette ribonucléase à double activité et montré que les deux activités sont catalysées par un même site actif.
La structure a également mis en évidence un site spécifique de liaison aux mononucléotides ce qui explique pourquoi l’activté 5’-3’ exonucléolytique est dépendante de l’état 5’ monophosphorylé ou 5’ hydroxyl du substrat qui est généralement le produit d’un clivage endonucléolytique.
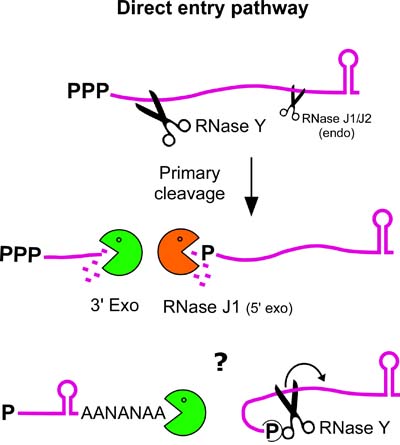
Des analyses du transcriptome et protéome de mutants des RNases J et Y ont confirmé l’importance globale de ces RNases chez B. subtilis. Nous avons observé qu’une déplétion de la RNase Y augmente la stabilité des ARNm totaux par un facteur supérieur à 2. Nous favorisons l’hypothèse que la voie principale de l’initiation de la dégradation de l’ARNm chez B. subtilis n’est pas exonucléolytique mais endonucléolytique.
Ceci suggère que la maturation et la dégradation de l’ARNm pourrait être plus similaire entre les organismes à Gram positif et ceux à Gram négatif que les modèles récents le laissaient supposer. Dans ce modèle l’activité 5’-3’ exonucléolytique de la RNase J ne servirait essentiellement qu’à dégrader des fragments d’ARN, produits d’un clivage endonucléolytique et protégés contre une dégradation exonucléolytique en 3’ par des structures secondaires (i.e. terminateurs).
Nous voulons maintenant mieux comprendre le rôle des RNases J et Y dans la régulation de gènes spécifiques chez B. subtilis afin d’identifier de nouvelles stratégies de contrôle de l’expression génique basées sur la stabilité des ARNm. L’étude des fonctions respectives de ces ribonucléases ainsi que celle de la régulation de leur expression va contribuer à mieux comprendre le métabolisme de l’ARNm chez les Bacilli.
Le comportement social des communautés bactériennes permet la survie et la dissémination dans divers habitats. La migration en masse (« swarming ») sur une surface nutritive précède la formation d’un biofilm classique et peut être considérée comme un mécanisme d’expansion territoriale. La migration dendritique de B. subtilis a lieu pendant plusieurs heures sous forme d’une monocouche cellulaire. Ce phenomène est mis à profit pour effectuer des analyses quantitatives spatio-temporelles de l’expression génique en cellule unique. Il nous permet d’analyser la localisation de l’ARNm et des RNases impliquées dans son métabolisme. Ce système élargit nos études de l’expression génique en faisant appel à un processus qui reflète plus fidèlement le comportement des bactéries dans la nature.
L’expression des aminoacyl-ARNt synthétases chez B. subtilis est également contrôlée par un mécanisme qui n’est pas observé chez E. coli: l’antiterminaison transcriptionnelle ARNt dépendante. Nos études de ces divers mécanismes de régulation ont de plus révélé l’importance de clivages stratégiques de l’ARN messager qui peuvent directement influencer l’expression des gènes. Ceci nous a amené, au cours des dernières années, à nous intéresser aux enzymes qui effectuent ces clivages.
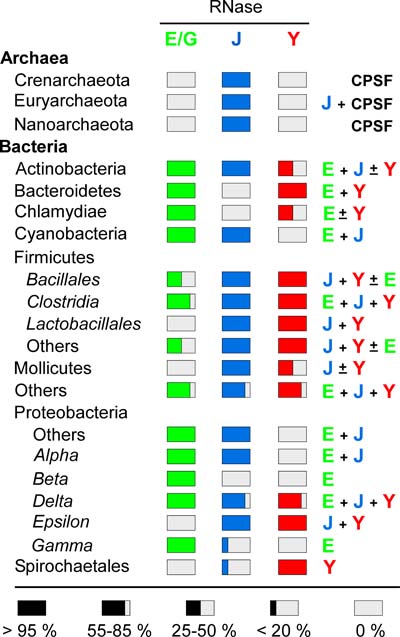
La RNase E est la ribonucléase majeure qui initie la dégradation des ARNm chez E. coli mais B. subtilis ne possède pas d’orthologue de cette enzyme clé. En étudiant le devenir de divers ARNm nous avons pu identifier deux nouvelles ribonucléases, les RNases J1/J2 et la RNase Y. De façon surprenante nous avons pu mettre en évidence que les ribonucléases J et Y ont une spécificité de clivage endonucléolytique similaire à celle de la RNase E d’E. coli et ce malgré une absence totale de similarité de séquence entre ces trois enzymes. Ainsi cette activité enzymatique a été inventée au moins trois fois de façon indépendante, illustrant un cas impressionnant d’évolution convergente. Il est probable que les trois RNases E, J et Y représentent les enzymes clé du métabolisme de l’ARN chez les eubactéries. En effet, toutes les espèces d’eubactéries possèdent au moins une de ces trois enzymes et bon nombre d’entre elles ont même les trois.